The third annual Targeted Protein Degradation (TPD) Symposium just wrapped up last month. It was kicked off with Poncho Meisenheimer, VP of Research and Development at Promega, likening the gathering of researchers to “kids in a biology candy store.” This playful analogy captured the vibrant energy and sense of exploration among the attendees, who convened to delve into the future possibilities of proximity-induced degradation. Poncho left attendees with three key questions to consider throughout the symposium:
How can we focus on quantitative measures of cellular events in relevant models?
How do we generate results that serve both human and AI models?
How do we best embrace the excitement of discovery?
Nearly 150 participants from both industry and academia attended the two-day symposium. It was held on September 11th and 12th at Promega’s R&D hub, the Kornberg Center, in Madison, Wisconsin. The event, now in its third year, provided a familiar environment where collaborations flourished, and many attendees rekindled connections forged through previous interactions or partnerships in the field.
In the opening remarks of our second annual Targeted Protein Degradation Symposium, Tom Livelli, VP of Life Sciences Products & Services at Promega, posed a question to the attendees: “What do you want to be able to do today that you can’t?” This aspirational question set the tone for an event where building connections to advance the study and application of proximity-induced degradation took center stage.
More than 90 attendees from academia and industry gathered September 20–21 for the two-day symposium, which was hosted in our inspirational Kornberg Center—the R&D heart of Promega. Through engaging talks, a poster session, “Learn n’ Burn” challenges and social gatherings, participants had the opportunity to reinforce existing collaborations and to connect with others who are making an impact in the field of targeted protein degradation.
Traditional approaches for protein degrader compound screening like Western blotting can be laborious, time consuming and cannot be streamlined with automation. By implementing a high-throughput, automated workflow that uses our CRISPER/Cas9 knock-in cell lines, live-cell bioluminescent assays and sensitive GloMax® Discover microplate readers, our custom assay services offer protein degradation profiling at an accelerated rate.
To do this, we collaborated with HighRes® Biosolutions, to develop an automated system that can screen up to 100 384-well plates each day, generating roughly 40,000 data points with minimal hands-on work.
An important step of building this system is to integrate four GloMax® Discover microplate readers into the automated system using instrument’s built-in SiLA2 communication driver. The driver software makes it easy to connect the microplate readers with HighRes® Biosolution’s robotic components.
While PROTACs might not be the topic of conversation at high society cocktail parties, or merit cover stories in glamor magazines, they’re certainly shaking up the drug discovery industry. PROTAC® degraders, together with related compounds like molecular glues and LYTACs, are the basic tools for a targeted protein degradation strategy. Research in this field is advancing rapidly, enabling the development of therapies for disease targets disease targets previously thought to be “undruggable”. This blog post provides an overview of PROTACs based on frequently asked questions.
With use and time things wear out. Tires get worn on a car, and you have the old tires removed, recycled, and replaced with new ones. Sometimes a part or piece of something isn’t made properly. For instance, if you are assembling a piece of furniture and you find a screw with no threads, you throw it out and get a screw that was made properly. The same thing holds true for cells. Components wear out (like tires) or get improperly made (a screw with no threads), or they simply have a limited lifetime so that they are available in the cell only when needed. These used and worn components need to be removed from the cell. One system that allows cells to recycle components and remove old or improperly functioning proteins is the Ubiquitin-Proteasome System (UPS). The UPS system relies on a series of small peptide tags, ubiquitin, to mark a protein for degradation. Researchers are now harnessing the UPS to target aberrant proteins in diseased cells through PROteolysis TArgeting Chimeras or PROTACs. PROTACs hold promise as highly efficacious therapeutics that can be directed to eliminate only a single protein. To take full advantage of the power of PROTACs, researchers need to understand the molecular underpinnings that are responsible for successful protein degradation. Here we review a paper that seeks to develop a computer model for predicting whether PROTAC ternary complex formation leads to ubiquitination and successful degradation of a target protein.
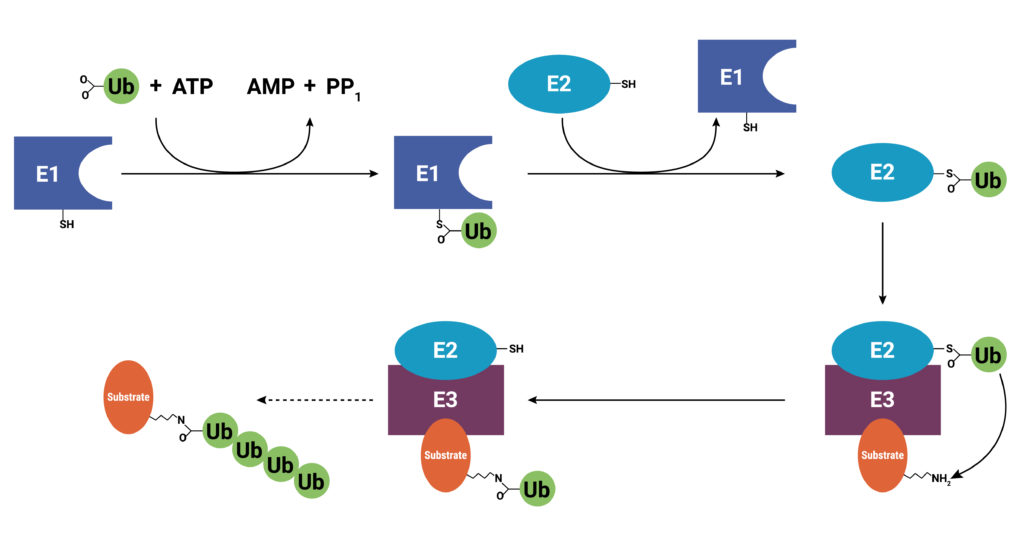
Proteins are targeted for degradation by the proteasome. A small chain of ubiquitin peptides (Ub) is added to available lysine residues of the target protein through the actions of three enzymes: E1, ubiquitin-activating enzyme; E2, ubiquitin-conjugating enzyme; and E3 ubiquitin ligase. After the addition of the Ub chain, the proteasome is recruited and the protein degraded.
Addressing the Intractable Target
Research to understand diseases including cancers, neurodegeneration, and auto-immune conditions has revealed that in many disease states, affected cells produce growth factors or enzymes that are constitutively active (“always on”). These proteins are targets for small molecule inhibitors that bind specific sites preventing the constitutive activity or signaling. More recently, biologics, or protein-based therapeutics, including monoclonal antibodies (mAb), have been developed that can bind and block inappropriate signaling pathways, especially those that allow cancer cells to escape immune system surveillance.
Unfortunately, up to 85% of targets have proven intractable to small molecule inhibitors, or they are not suitable for a biologics approach. Oftentimes, the target protein doesn’t have a great place to bind a small molecule, so even though inhibitors might exist they cannot bind well enough to be effective. Or, as in the case of many cancers, the diseased cell manages to overcome the effect of the inhibitor by overexpressing the target. Still other aberrant proteins associated with diseases haven’t gained function to cause a disease; they have instead, lost function, so designing an inhibitor of the protein is not a workable strategy. Enter the PROTAC.
G protein-coupled receptors (GPCRs) comprise a large group of cell surface receptors, characterized by the unique structural property of crossing the cell membrane seven times. They respond to a diverse group of signaling molecules, such as peptides, neurotransmitters, cytokines, hormones and other small molecules (1). Upon activation, GPCRs interact with GTP-binding (G) proteins and arrestins to regulate a wide variety of signaling pathways. This broad range of functions makes GPCRs attractive targets for drug discovery. The importance of GPCR research was highlighted in 2012, with the Nobel Prize in chemistry being awarded to Robert Lefkowitz and Brian Kobilka “for studies of G-protein–coupled receptors”.
Based on structure and function, GPCRs are categorized into six classes, A–F. The class A GPCRs, or rhodopsin-like receptors, have been studied extensively due to their association with many types of diseases (2). Within the class A GPCRs is a group that share a highly conserved structural motif (3) and respond to chemokines—small “chemotactic cytokines” that stimulate cell migration, especially that of white blood cells (4). A subfamily of class A GPCRs respond to chemokines that have two cysteine residues near the N-terminus, known as CC chemokines. GPCRs activated by CC chemokines are called CC chemokine receptors or CCRs, and these interactions have been implicated in both pro- and anti-cancer pathways (5).
In 1963, Jennifer Harvey was studying Moloney murine leukemia virus (MMLV) at the cancer research department of the London Hospital Research Laboratories. After routine transfers of plasma from MMLV-infected rats to mice, she made an unusual discovery. In addition to the expected leukemia, the mice that received the plasma developed solid tumors (soft-tissue sarcomas), primarily in the spleen (1). A few years later, Werner Kirsten at the University of Chicago observed similar results working with mouse erythroblastosis virus (MEV) (2).
Subsequent research, with the advent of genome sequencing, showed that a cellular rat gene had been incorporated into the viral genome in both cases (3). These genomic sequences contained a mutation later shown to be responsible for the development of sarcomas, and the word “oncogene” became a common part of the vocabulary in cancer publications during the early 1980s (4). Harvey’s discovery led to the naming of the corresponding rat sarcoma oncogene as HRAS, while Kirsten’s related oncogene was named KRAS. Several laboratories, working independently, cloned the human homolog of the viral HRAS gene in 1982 (3). The human KRAS gene was cloned shortly thereafter, as well as a third RAS gene, named NRAS (3). Additional studies showed that a single point mutation in each of these genes led to oncogenic activation, and they have been popular targets for anticancer drug discovery efforts ever since.
The Dana-Farber Targeted Protein Degradation Webinar Series discusses new discoveries and modalities in protein degradation.
In this webinar, Senior Research Scientist, Dr. Danette Daniels, focuses primarily on proteolysis-targeting chimeras, or PROTACs. A variety of topics are covered including the design, potency, and efficacy of PROTACs in targeted protein degradation. Watch the video below to learn more about how PROTACs are shifting perspectives through fascinating research and discoveries in targeted protein degradation.
Learn more about targeted protein degradation and PROTACS here.
Transcriptional activation of genes within the nucleus of eukaryotic cells occurs by a variety of mechanisms. Typically, these mechanisms rely on the interaction of regulatory proteins (transcriptional activators or repressors) with specific DNA sequences that control gene expression. Upon DNA binding, regulatory proteins also interact with other proteins that are part of the RNA polymerase II transcriptional complex.
One type of transcriptional activation relies on inducing a conformational change in chromatin, the DNA-protein complex that makes up each chromosome within a cell. In a broad sense, “extended” or loosely wound chromatin is more accessible to transcription factors and can signify an actively transcribed gene. In contrast, “condensed” chromatin hinders access to transcription factors and is characteristic of a transcriptionally inactive state. Acetylation of lysine residues in histones—the primary constituents of the chromatin backbone—results in opening up the chromatin and consequent gene activation. Disruption of histone acetylation pathways is implicated in many types of cancer (1).
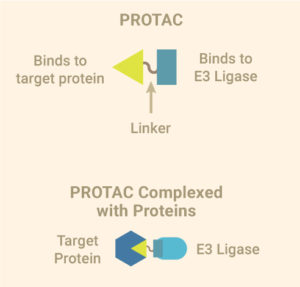
An illustration of PROTAC structure and the proteins it binds.
Targeting a single protein and making it disappear from the cell is quite the magic trick, and there are various molecular tools available for this task. You can use RNA interference, which prevents a protein from being made, inhibitors that bind the protein, rendering it unavailable for use or even gene editing tools like CRISPR that can remove it from the genome. But did you know that you can target an existing protein for destruction, using the cell’s own garbage disposal system to degrade the protein? All you need is a molecule that can connect your protein to one with a role in cellular protein degradation and your protein can be destroyed.
XWe use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To learn more about our approach to Privacy we invite you to Read More
By clicking “Accept All”, you consent to the use of ALL the cookies. However you may visit Cookie Settings to provide a controlled consent.
We use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To find out more about cookies and how to manage cookies, read our Cookie Policy.
If you are located in the EEA, the United Kingdom, or Switzerland, you can change your settings at any time by clicking Manage Cookie Consent in the footer of our website.
Necessary cookies are absolutely essential for the website to function properly. These cookies ensure basic functionalities and security features of the website, anonymously.
Cookie
Duration
Description
cookielawinfo-checbox-analytics
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Analytics".
cookielawinfo-checbox-functional
11 months
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Functional".
cookielawinfo-checbox-others
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Other.
cookielawinfo-checkbox-advertisement
1 year
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Advertisement".
cookielawinfo-checkbox-necessary
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookies is used to store the user consent for the cookies in the category "Necessary".
cookielawinfo-checkbox-performance
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Performance".
gdpr_status
6 months 2 days
This cookie is set by the provider Media.net. This cookie is used to check the status whether the user has accepted the cookie consent box. It also helps in not showing the cookie consent box upon re-entry to the website.
lang
This cookie is used to store the language preferences of a user to serve up content in that stored language the next time user visit the website.
viewed_cookie_policy
11 months
The cookie is set by the GDPR Cookie Consent plugin and is used to store whether or not user has consented to the use of cookies. It does not store any personal data.
Analytical cookies are used to understand how visitors interact with the website. These cookies help provide information on metrics the number of visitors, bounce rate, traffic source, etc.
Cookie
Duration
Description
SC_ANALYTICS_GLOBAL_COOKIE
10 years
This cookie is associated with Sitecore content and personalization. This cookie is used to identify the repeat visit from a single user. Sitecore will send a persistent session cookie to the web client.
vuid
2 years
This domain of this cookie is owned by Vimeo. This cookie is used by vimeo to collect tracking information. It sets a unique ID to embed videos to the website.
WMF-Last-Access
1 month 18 hours 24 minutes
This cookie is used to calculate unique devices accessing the website.
_ga
2 years
This cookie is installed by Google Analytics. The cookie is used to calculate visitor, session, campaign data and keep track of site usage for the site's analytics report. The cookies store information anonymously and assign a randomly generated number to identify unique visitors.
_gid
1 day
This cookie is installed by Google Analytics. The cookie is used to store information of how visitors use a website and helps in creating an analytics report of how the website is doing. The data collected including the number visitors, the source where they have come from, and the pages visted in an anonymous form.
Advertisement cookies are used to provide visitors with relevant ads and marketing campaigns. These cookies track visitors across websites and collect information to provide customized ads.
Cookie
Duration
Description
IDE
1 year 24 days
Used by Google DoubleClick and stores information about how the user uses the website and any other advertisement before visiting the website. This is used to present users with ads that are relevant to them according to the user profile.
test_cookie
15 minutes
This cookie is set by doubleclick.net. The purpose of the cookie is to determine if the user's browser supports cookies.
VISITOR_INFO1_LIVE
5 months 27 days
This cookie is set by Youtube. Used to track the information of the embedded YouTube videos on a website.
Performance cookies are used to understand and analyze the key performance indexes of the website which helps in delivering a better user experience for the visitors.
Cookie
Duration
Description
YSC
session
This cookies is set by Youtube and is used to track the views of embedded videos.
_gat_UA-62336821-1
1 minute
This is a pattern type cookie set by Google Analytics, where the pattern element on the name contains the unique identity number of the account or website it relates to. It appears to be a variation of the _gat cookie which is used to limit the amount of data recorded by Google on high traffic volume websites.